InVivoMAb anti-mouse LPAM-1 (Integrin α4β7)
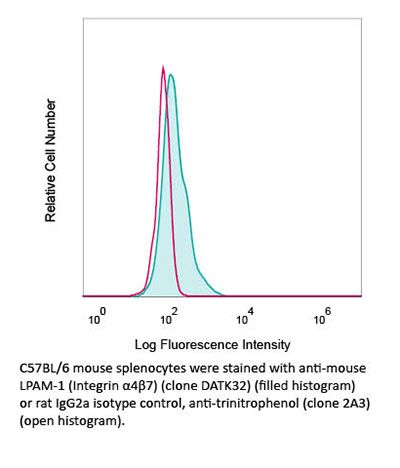
Product Description
Specifications
| Isotype | Rat IgG2a, κ |
|---|---|
| Recommended Isotype Control(s) | InVivoMAb rat IgG2a isotype control, anti-trinitrophenol |
| Recommended Dilution Buffer | InVivoPure pH 7.0 Dilution Buffer |
| Conjugation | This product is unconjugated. Conjugation is available via our Antibody Conjugation Services. |
| Immunogen | TK1 cells |
| Reported Applications |
in vivo Integrin α4β7 neutralization Flow cytometry |
| Formulation |
PBS, pH 7.0 Contains no stabilizers or preservatives |
| Endotoxin |
≤1EU/mg (≤0.001EU/μg) Determined by LAL assay |
| Purity |
≥95% Determined by SDS-PAGE |
| Sterility | 0.2 µm filtration |
| Production | Purified from cell culture supernatant in an animal-free facility |
| Purification | Protein G |
| RRID | AB_1107713 |
| Molecular Weight | 150 kDa |
| Storage | The antibody solution should be stored at the stock concentration at 4°C. Do not freeze. |
| Need a Custom Formulation? | See All Antibody Customization Options |
Application References
-
Duc, D., et al (2019). "Disrupting Myelin-Specific Th17 Cell Gut Homing Confers Protection in an Adoptive Transfer Experimental Autoimmune Encephalomyelitis" Cell Rep 29(2): 378-390.e374.
PubMed
Multiple sclerosis (MS) is a common autoimmune disease of the CNS. Although an association between MS and inflammatory bowel diseases is observed, the link connecting intestinal immune responses and neuroinflammation remains unclear. Here we show that encephalitogenic Th17 cells infiltrate the colonic lamina propria before neurological symptom development in two murine MS models, active and adoptive transfer experimental autoimmune encephalomyelitis (EAE). Specifically targeting Th17 cell intestinal homing by blocking the α4β7-integrin and its ligand MAdCAM-1 pathway impairs T cell migration to the large intestine and dampens EAE severity in the Th17 cell adoptive transfer model. Mechanistically, myelin-specific Th17 cells proliferate in the colon and affect gut microbiota composition. The beneficial effect of blocking the α4β7-integrin and its ligand MAdCAM-1 pathway on EAE is interdependent with gut microbiota. Those results show that disrupting myelin-specific Th17 cell trafficking to the large intestine harnesses neuroinflammation and suggests that the gut environment and microbiota catalyze the encephalitogenic properties of Th17 cells.
-
Bemark, M., et al (2016). "Limited clonal relatedness between gut IgA plasma cells and memory B cells after oral immunization" Nat Commun 7: 12698.
PubMed
Understanding how memory B cells are induced and relate to long-lived plasma cells is important for vaccine development. Immunity to oral vaccines has been considered short-lived because of a poor ability to develop IgA B-cell memory. Here we demonstrate that long-lived mucosal IgA memory is readily achieved by oral but not systemic immunization in mouse models with NP hapten conjugated with cholera toxin and transfer of B1-8(high)/GFP(+) NP-specific B cells. Unexpectedly, memory B cells are poorly related to long-lived plasma cells and less affinity-matured. They are α4β7-integrin(+)CD73(+)PD-L2(+)CD80(+) and at systemic sites mostly IgM(+), while 80% are IgA(+) in Peyer’s patches. On reactivation, most memory B cells in Peyer’s patches are GL7(-), but expand in germinal centres and acquire higher affinity and more mutations, demonstrating strong clonal selection. CCR9 expression is found only in Peyer’s patches and appears critical for gut homing. Thus, gut mucosal memory possesses unique features not seen after systemic immunization.
-
Rosser, E. C., et al (2014). "Regulatory B cells are induced by gut microbiota-driven interleukin-1beta and interleukin-6 production" Nat Med 20(11): 1334-1339.
PubMed
Regulatory B cells (Breg cells) differentiate in response to inflammation and subsequently restrain excessive immune responses via the release of interleukin-10 (IL-10). However, the precise inflammatory signals governing their differentiation remain to be elucidated. Here we show that the gut microbiota promotes the differentiation of Breg cells in the spleen as well as in the mesenteric lymph nodes. Perturbation of the gut microbiome imposed either by antibiotic treatment or by changes in the sterility of housing conditions reduces the number and function of Breg cells. Following the induction of arthritis, IL-1beta and IL-6 are produced only in conventionally housed mice and both cytokines directly promote Breg cell differentiation and IL-10 production. Mice lacking IL-6 receptor (IL-6R) or IL-1 receptor 1 (IL-1R1) specifically on B cells have a reduced number of IL-10-producing B cells and develop exacerbated arthritis compared to control animals. Thus, in response to inflammatory signals induced by both the gut flora and arthritis, Breg cells increase in number and restrain excessive inflammation.
-
Sheridan, B. S., et al (2014). "Oral infection drives a distinct population of intestinal resident memory CD8(+) T cells with enhanced protective function" Immunity 40(5): 747-757.
PubMed
The intestinal mucosa promotes T cell responses that might be beneficial for effective mucosal vaccines. However, intestinal resident memory T (Trm) cell formation and function are poorly understood. We found that oral infection with Listeria monocytogenes induced a robust intestinal CD8 T cell response and blocking effector T cell migration showed that intestinal Trm cells were critical for secondary protection. Intestinal effector CD8 T cells were predominately composed of memory precursor effector cells (MPECs) that rapidly upregulated CD103, which was needed for T cell accumulation in the intestinal epithelium. CD103 expression, rapid MPEC formation, and maintenance in intestinal tissues were dependent on T cell intrinsic transforming growth factor beta signals. Moreover, intestinal Trm cells generated after intranasal or intravenous infection were less robust and phenotypically distinct from Trm cells generated after oral infection, demonstrating the critical contribution of infection route for directing the generation of protective intestinal Trm cells.
Product Citations
-
Colonic inflammation triggers β cell proliferation during obesity development via a liver-to-pancreas interorgan mechanism.
In JCI Insight on 8 May 2025 by Kubo, H., Imai, J., et al.
PubMed
Under insulin-resistant conditions, such as obesity, pancreatic β cells adaptively proliferate and secrete more insulin to prevent blood glucose elevation. We previously reported hepatic ERK activation during obesity development to stimulate a neuronal relay system, consisting of afferent splanchnic nerves from the liver and efferent vagal nerves to the pancreas, thereby triggering adaptive β cell proliferation. However, the mechanism linking obesity with the interorgan system originating in hepatic ERK activation remains unclear. Herein, we clarified that colonic inflammation promotes β cell proliferation through this interorgan system from the liver to the pancreas. First, dextran sodium sulfate (DSS) treatment induced colonic inflammation and hepatic ERK activation as well as β cell proliferation, all of which were suppressed by blockades of the neuronal relay system by several approaches. In addition, treatment with anti-lymphocyte Peyer's patch adhesion molecule-1 (anti-LPAM1) antibody suppressed β cell proliferation induced by DSS treatment. Importantly, high-fat diet (HFD) feeding also elicited colonic inflammation, and its inhibition by anti-LPAM1 antibody administration suppressed hepatic ERK activation and β cell proliferation induced by HFD. Thus, colonic inflammation triggers adaptive β cell proliferation via the interorgan mechanism originating in hepatic ERK activation. The present study revealed a potentially novel role of the gastrointestinal tract in the maintenance of β cell regulation.
-
Effect of Integrin Blockade on Experimental Spondyloarthritis.
In Biomolecules on 31 October 2024 by Yau, E., Lim, M., et al.
PubMed
Spondyloarthritis (SpA) describes a group of diseases characterized by chronic inflammation in the spine and peripheral joints. While pathogenesis is still unclear, proinflammatory gut-derived immune cells have been identified in the joints of SpA patients. We previously identified an enriched population of integrin-expressing cells in the joints of SpA patients. Entry of gut-derived cells into joints may be mediated by these integrins. In the current study, we used the SKG murine model of SpA to study the impact of integrin blockade. Mice were injected with antibodies against the integrin α4β7 or the β7 monomer twice a week. Treatment with antibodies against α4β7 reduced disease severity in curdlan-injected SKG mice, with disease scores being comparable between treatment initiation times. Targeting the β7 monomer led to reduced arthritis severity compared to targeting the α4β7 dimer. Treatment with antibodies against α4β7 or β7 decreased expression of these integrins in CD4+ T cells, with the frequency of αE+β7+ T cells in the spleen and lymph nodes correlating with disease severity. In summary, we showed that integrin blockade showed potential for ameliorating disease in a murine model of SpA, lending support for further studies testing integrin blockade in SpA.
-
Overcoming Resistance to Immune Checkpoint Inhibitor Therapy Using Calreticulin-Inducing Nanoparticle.
In Pharmaceutics on 9 June 2023 by Chandrasekar, S. V., Singh, A., et al.
PubMed
Nanoparticles (NPs) have the ability to transform poorly immunogenic tumors into activated 'hot' targets. In this study, we investigated the potential of a liposome-based nanoparticle (CRT-NP) expressing calreticulin as an in-situ vaccine to restore sensitivity to anti-CTLA4 immune checkpoint inhibitor (ICI) in CT26 colon tumors. We found that a CRT-NP with a hydrodynamic diameter of approximately 300 nm and a zeta potential of approximately +20 mV induced immunogenic cell death (ICD) in CT-26 cells in a dose-dependent manner. In the mouse model of CT26 xenograft tumors, both CRT-NP and ICI monotherapy caused moderate reductions in tumor growth compared to the untreated control group. However, the combination therapy of CRT-NP and anti-CTLA4 ICI resulted in remarkable suppression of tumor growth rates (>70%) compared to untreated mice. This combination therapy also reshaped the tumor microenvironment (TME), achieving the increased infiltration of antigen-presenting cells (APCs) such as dendritic cells and M1 macrophages, as well as an abundance of T cells expressing granzyme B and a reduction in the population of CD4+ Foxp3 regulatory cells. Our findings indicate that CRT-NPs can effectively reverse immune resistance to anti-CTLA4 ICI therapy in mice, thereby improving the immunotherapeutic outcome in the mouse model.
-
The Appendix Orchestrates T-Cell Mediated Immunosurveillance in Colitis-Associated Cancer.
In Cell Mol Gastroenterol Hepatol on 5 November 2022 by Collard, M. K., Tourneur-Marsille, J., et al.
PubMed
Although appendectomy may reduce colorectal inflammation in patients with ulcerative colitis (UC), this surgical procedure has been suggested to be associated with an increased risk of colitis-associated cancer (CAC). Our aim was to explore the mechanism underlying the appendectomy-associated increased risk of CAC.