InVivoMAb anti-mouse L-Selectin (CD62L)
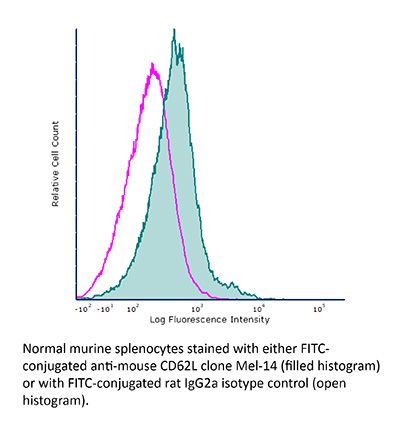
Product Description
Specifications
| Isotype | Rat IgG2a, κ |
|---|---|
| Recommended Isotype Control(s) | InVivoMAb rat IgG2a isotype control, anti-trinitrophenol |
| Recommended Dilution Buffer | InVivoPure pH 7.0 Dilution Buffer |
| Conjugation | This product is unconjugated. Conjugation is available via our Antibody Conjugation Services. |
| Immunogen | C3H/eb mouse B lymphoma 38C-13 |
| Reported Applications | in vivo CD62L neutralization |
| Formulation |
PBS, pH 7.0 Contains no stabilizers or preservatives |
| Endotoxin |
≤1EU/mg (≤0.001EU/μg) Determined by LAL assay |
| Purity |
≥95% Determined by SDS-PAGE |
| Sterility | 0.2 µm filtration |
| Production | Purified from cell culture supernatant in an animal-free facility |
| Purification | Protein G |
| RRID | AB_1107665 |
| Molecular Weight | 150 kDa |
| Storage | The antibody solution should be stored at the stock concentration at 4°C. Do not freeze. |
| Need a Custom Formulation? | See All Antibody Customization Options |
Application References
-
Guidotti, L. G., et al (2015). "Immunosurveillance of the liver by intravascular effector CD8(+) T cells" Cell 161(3): 486-500.
PubMed
Effector CD8(+) T cells (CD8 TE) play a key role during hepatotropic viral infections. Here, we used advanced imaging in mouse models of hepatitis B virus (HBV) pathogenesis to understand the mechanisms whereby these cells home to the liver, recognize antigens, and deploy effector functions. We show that circulating CD8 TE arrest within liver sinusoids by docking onto platelets previously adhered to sinusoidal hyaluronan via CD44. After the initial arrest, CD8 TE actively crawl along liver sinusoids and probe sub-sinusoidal hepatocytes for the presence of antigens by extending cytoplasmic protrusions through endothelial fenestrae. Hepatocellular antigen recognition triggers effector functions in a diapedesis-independent manner and is inhibited by the processes of sinusoidal defenestration and capillarization that characterize liver fibrosis. These findings reveal the dynamic behavior whereby CD8 TE control hepatotropic pathogens and suggest how liver fibrosis might reduce CD8 TE immune surveillance toward infected or transformed hepatocytes.
-
Brinkman, C. C., et al (2016). "Treg engage lymphotoxin beta receptor for afferent lymphatic transendothelial migration" Nat Commun 7: 12021.
PubMed
Regulatory T cells (Tregs) are essential to suppress unwanted immunity or inflammation. After islet allo-transplant Tregs must migrate from blood to allograft, then via afferent lymphatics to draining LN to protect allografts. Here we show that Tregs but not non-Treg T cells use lymphotoxin (LT) during migration from allograft to draining LN, and that LT deficiency or blockade prevents normal migration and allograft protection. Treg LTalphabeta rapidly modulates cytoskeletal and membrane structure of lymphatic endothelial cells; dependent on VCAM-1 and non-canonical NFkappaB signalling via LTbetaR. These results demonstrate a form of T-cell migration used only by Treg in tissues that serves an important role in their suppressive function and is a unique therapeutic focus for modulating suppression.
-
Cremasco, V., et al (2014). "B cell homeostasis and follicle confines are governed by fibroblastic reticular cells" Nat Immunol 15(10): 973-981.
PubMed
Fibroblastic reticular cells (FRCs) are known to inhabit T cell-rich areas of lymphoid organs, where they function to facilitate interactions between T cells and dendritic cells. However, in vivo manipulation of FRCs has been limited by a dearth of genetic tools that target this lineage. Here, using a mouse model to conditionally ablate FRCs, we demonstrated their indispensable role in antiviral T cell responses. Unexpectedly, loss of FRCs also attenuated humoral immunity due to impaired B cell viability and follicular organization. Follicle-resident FRCs established a favorable niche for B lymphocytes via production of the cytokine BAFF. Thus, our study indicates that adaptive immunity requires an intact FRC network and identifies a subset of FRCs that control B cell homeostasis and follicle identity.
-
Harp, J. R., et al (2010). "Memory T cells are enriched in lymph nodes of selectin-ligand-deficient mice" J Immunol 185(10): 5751-5761.
PubMed
Fucosyltransferase-IV and -VII double knockout (FtDKO) mice reveal profound impairment in T cell trafficking to lymph nodes (LNs) due to an inability to synthesize selectin ligands. We observed an increase in the proportion of memory/effector (CD44(high)) T cells in LNs of FtDKO mice. We infected FtDKO mice with lymphocytic choriomeningitis virus to generate and track Ag-specific CD44(high)CD8 T cells in secondary lymphoid organs. Although frequencies were similar, total Ag-specific effector CD44(high)CD8 T cells were significantly reduced in LNs, but not blood, of FtDKO mice at day 8. In contrast, frequencies of Ag-specific memory CD44(high)CD8 T cells were up to 8-fold higher in LNs of FtDKO mice at day 60. Because wild-type mice treated with anti-CD62L treatment also showed increased frequencies of CD44(high) T cells in LNs, we hypothesized that memory T cells were preferentially retained in, or preferentially migrated to, FtDKO LNs. We analyzed T cell entry and egress in LNs using adoptive transfer of bone fide naive or memory T cells. Memory T cells were not retained longer in LNs compared with naive T cells; however, T cell exit slowed significantly as T cell numbers declined. Memory T cells were profoundly impaired in entering LNs of FtDKO mice; however, memory T cells exhibited greater homeostatic proliferation in FtDKO mice. These results suggest that memory T cells are enriched in LNs with T cell deficits by several mechanisms, including longer T cell retention and increased homeostatic proliferation.
Product Citations
-
Enantiomer-dependent and modification-free DNA matrix as an adjuvant for subunit vaccines against SARS-CoV-2 or pneumococcal infections.
In Nat Biomed Eng on 1 December 2025 by Li, C., Li, Y., et al.
PubMed
The emergence of novel infectious disease has intensified demand for more advanced vaccine development and more potent adjuvants to enhance immunogenicity. Here we introduce a dynamic DNA supramolecular matrix assembled from five unmodified, short DNA single strands, serving as a safe, multifaceted adjuvant platform. This DNA matrix elicits a robust humoral response with minimal adverse effects, generating potent neutralizing antibodies and conferring robust protection against SARS-CoV-2 and Streptococcus pneumoniae infections. Its dynamic colloidal feature prolongs the in vivo retention of both DNA and antigen, facilitating lymphatic-targeted transportation and presentation. This process leads to a robust pro-inflammatory response in both the vaccinated site and draining lymph node, which, in turn, promotes the recruitment and activation of immune cells, leading to a rapid, effective antigen-specific antibody response. The enhanced function of DNA matrix depends on the canonical TLR9-MyD88 signalling axis in dendritic cells. In addition, only right-handed, not left-handed, chirality of the DNA strands forms D-DNA matrix and promotes immune activations. Thus, this DNA matrix functions as an all-in-one adjuvant platform, opening promising avenues for future vaccine design.
-
CD301b+ dendritic cell-derived IL-2 dictates CD4+ T helper cell differentiation.
In Nat Commun on 26 February 2025 by Tatsumi, N., El-Fenej, J., et al.
PubMed
T helper (Th) cell differentiation is fundamental to functional adaptive immunity. Different subsets of dendritic cells (DC) preferentially induce different types of Th cells, but the DC-derived mechanism for Th type 2 (Th2) differentiation is not fully understood. Here, we show that in mice, CD301b+ DCs, a major Th2-inducing DC subset, drive Th2 differentiation through cognate interaction by rapidly inducing IL-2 receptor signalling in CD4+ T cells. Mechanistically, CD40 engagement prompts IL-2 production selectively from CD301b+ DCs to maximize CD25 expression in CD4+ T cells, which instructs the Th2 fate decision, while simultaneously skewing CD4+ T cells away from the T follicular helper fate. Moreover, CD301b+ DCs utilize their own CD25 to facilitate directed action of IL-2 toward cognate CD4+ T cells, as genetic deletion of CD25 in CD301b+ DCs results in reduced IL-2-mediated signalling in antigen-specific CD4+ T cells and hence their Th2 differentiation. These results highlight the critical role of DC-intrinsic CD40-IL-2 axis in Th cell fate decision.
-
A lymphoid tissue chemokine checkpoint prevents loss of CD8+T cell functionality
In bioRxiv on 22 September 2024 by Altenburger, L. M., Claudino Carvoeiro, D., et al.
-
Therapeutic potential of co-signaling receptor modulation in hepatitis B.
In Cell on 25 July 2024 by Andreata, F., Laura, C., et al.
PubMed
Reversing CD8+ T cell dysfunction is crucial in treating chronic hepatitis B virus (HBV) infection, yet specific molecular targets remain unclear. Our study analyzed co-signaling receptors during hepatocellular priming and traced the trajectory and fate of dysfunctional HBV-specific CD8+ T cells. Early on, these cells upregulate PD-1, CTLA-4, LAG-3, OX40, 4-1BB, and ICOS. While blocking co-inhibitory receptors had minimal effect, activating 4-1BB and OX40 converted them into antiviral effectors. Prolonged stimulation led to a self-renewing, long-lived, heterogeneous population with a unique transcriptional profile. This includes dysfunctional progenitor/stem-like (TSL) cells and two distinct dysfunctional tissue-resident memory (TRM) populations. While 4-1BB expression is ubiquitously maintained, OX40 expression is limited to TSL. In chronic settings, only 4-1BB stimulation conferred antiviral activity. In HBeAg+ chronic patients, 4-1BB activation showed the highest potential to rejuvenate dysfunctional CD8+ T cells. Targeting all dysfunctional T cells, rather than only stem-like precursors, holds promise for treating chronic HBV infection.